Propriedades e absorção da água
A água é uma das substâncias mais
importantes do planeta e sem água não há vida. A conquista do ambiente
terrestre pelas plantas se deu pelo desenvolvimento de raízes para mantê-las
eretas no solo, um sistema vascular avançado para absorver e transportar a água
e uma epiderme e estruturas especializadas (estômatos) para conservá-la.
A água constitui cerca de 80 a
90% do corpo de um vegetal metabolicamente ativo. Boa parte da água absorvida é
perdida para a atmosfera em forma de vapor –TRANSPIRAÇÃO. Devemos
saber que a água é um dos itens mais importantes e mais abundantes, porém é um
fator limitante com relação à diversidade e distribuição de espécies.
A absorção de água pelas células
gera, interiormente, uma força denominada TURGOR sendo
extremamente necessária em vários processos fisiológicos, como alongamento
celular, trocas gasosas nas folhas e transporte de solutos pelo floema. A falta
de água pode ser denominada como déficit hídrico, ou estresse hídrico e
desencadeia na planta vários processos para que ela continue com suas funções
vitais, como o fechamento estomático, a redução da fotossíntese e da respiração
e interfere em outros processos básicos.
A importância da água se deve ao
fato das características químicas da molécula, que conferem propriedades
físico-químicas, sendo considerada como o solvente universal. A água é composta
por três moléculas, sendo uma o oxigênio (O) ligado covalentemente às duas
outras moléculas que são de hidrogênio (H). Cada molécula da água possui
cargas: o oxigênio, mais eletronegativo que o hidrogênio, tende a atrair
elétrons para si, resultando em uma carga negativa parcial na extremidade da
molécula de oxigênio e uma carga positiva parcial em cada molécula de
hidrogênio. A molécula de água não possui carga líquida. A fraca atração
eletrostática da molécula é denominada de PONTE DE HIDROGÊNIO.
Essas duas moléculas quando separadas podem interagir com muitas outras
moléculas formando outras substâncias.
A água
possui propriedades físicas - Propriedade Solvente, Propriedade de Coesão e
Adesão e Propriedade Térmica – fazendo com que as moléculas de Hidrogênio e
Oxigênio, se movimentem através de três processos: fluxo de massa, difusão e
osmose.
1. Fluxo de massa: quando uma força externa é
aplicada, as moléculas tendem a se mover na mesma direção em massa. Ex.: água e
soluto movem-se através do xilema por fluxo em massa;
2. Difusão: movimento espontâneo (ao acaso)
de partículas, pela sua própria energia. Ex.: um sistema em contínua
movimentação;
3. Osmose: movimento da água entre
membranas semipermeáveis no sentido do equilíbrio de concentrações entre dois
meios.
Mas por
onde as plantas absorvem água?
As
plantas absorvem água do solo através das raízes. A zona de maior
absorção é conhecida como Zona Pilífera (como estudamos na
disciplina de Morfologia e Sistemática Vegetal), que contém tricomas (pelos)
finos que estão em contato íntimo com as partículas do solo. Além da importante
função de absorção de água as raízes desempenham a função de sustentação,
armazenamento de reservas, síntese de substâncias e absorção de nutrientes.

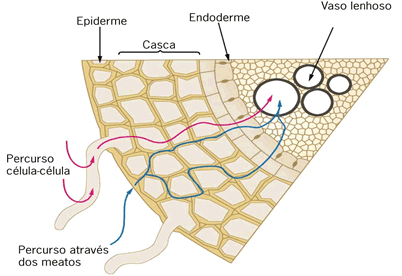
Após ser
absorvida pelas raízes, a água pode se movimentar por meio de três vias dentro
do xilema:
1. Via
Apoplasto: caminho contínuo da água pelas paredes e espaços celulares, isto
é, água não atravessa membrana. O movimento é extracelular;
2. Via
Transmembrana: caminho da água através de várias membranas. O movimento é
intracelular;
3. Via
Simplasto: caminho da água de uma célula a outra por meio de plasmodesmos.
A água atravessa a membrana, porém não o protoplasto.

Para
relembrarmos, o xilema, sistema de vasos condutores de água e
substâncias inorgânicas, é composto por dois tipos de células: traqueídes
e elementos de vaso. A ascensão da água pelo xilema é um processo que
requer uma força motriz bastante elevada (quanto mais alta for a planta, maior
deverá ser a força que permite a chegada da seiva xilemática até o ápice
caulinar).
A Teoria
de Dixon (1914) – Teoria da Coesão, Tensão e Adesão - é a mais
aceita universalmente para explicar como ocorre a ascensão da água e
substâncias inorgânicas. É “regida, basicamente, por um gradiente de
potencial hídrico (entre a atmosfera, a planta e o solo), pelas propriedades de
coesão e adesão das moléculas de água e pela força de tensão nos vasos
xilemáticos”.
Essa
Teoria é chamada de COESÃO-TENSÃO porque requer que as propriedades
coesivas da água sejam capazes de suportar a tensão da coluna de água
do xilema. A propriedade de adesão das moléculas da água às
paredes dos traqueídes e elementos de vaso é também importante para a ascensão
da seiva.

Transpiração
Transpiração
é a perda de água pelas plantas na forma de vapor
d’água. Cerca de 95% da água é perdida pela transpiração, sendo o restante
usado no metabolismo e no crescimento vegetal. A transpiração ocorre em
qualquer parte do organismo vegetal acima do solo. Pouca quantidade pode ser
perdida através de lenticelas (pequenas aberturas da casca de caules e ramos
jovens); porém, a maior proporção ocorre nas folhas.
Vamos
relembrar um pouco a anatomia foliar: há duas epidermes (adaxial
ou superior e abaxial ou inferior); acima da epiderme adaxial pode existir
ainda uma camada de cutícula que é uma barreira contra a perda
de água. No mesófilo das folhas, as células do parênquima paliçádico e
lacunoso conectam os espaços intercelulares, além de realizarem
fotossíntese. Ocorrem nas epidermes os estômatos, que têm função
primordial das trocas gasosas.
 |
| Figura 1 – anatomia foliar |
Legenda: 1) cutícula 2) epiderme
adaxial 3) parênquima paliçádico 4) parênquima
lacunoso 5) Epiderme abaxial 6) estômato 7) células
guardas 8) xilema 9)floema 10) tecidos
condutores do sistema vascular.
Na figura
2, podemos observar que os estômatos são compostos por uma
abertura, denominada ostíolo. Esta abertura é circundada por duas células-guardarepletas
de cloroplastos. Podem ocorrer, na maioria das plantas, células-subsidiárias ao
lado de cada uma das células-guarda e uma câmara subestomática.
 |
| Figura 2 – Estômato. Fonte: PERES, 2005, www.ciagri.usp.br, acesso em 26/06/2006 |
A Teoria mais
aceita de como funciona a abertura e o fechamento estomático foi proposta por Iamamura,
em 1943, que relata a existência de um fluxo de potássio (K+) no
interior das células-guarda, gerando um gradiente de potencial hídrico. É um
processo com gasto de energia, isto é, para que ocorra o fluxo de K+ para
o interior das células-guarda, ocorre ativação das bombas de prótons (H+)
localizadas nas membranas plasmáticas das células. Quando o conteúdo de K+aumenta
nas células-guarda, os estômatos abrem e, quando diminui a quantidade de K+ nas
células-guarda, os estômatos fecham.
Para que
o balanço de íons e prótons não seja prejudicado, já que ocorre uma intensa
entrada de potássio, que tem carga positiva, os ânions cloro (Cl-) emalato2- contribuem
para a neutralidade elétrica das células-guarda. A concentração desses ânions
aumenta na abertura dos estômatos e diminui quando o estômato se fecha.
Recentemente
os estudos sobre o papel da sacarose na osmorregulação das
células-guarda foram retomados. Esses estudos mostram que a abertura dos
estômatos pela manhã está relacionada com o aumento na concentração de K+ nas
células-guarda e, no período da tarde, quando os estômatos se fecham, há um
declínio no conteúdo de sacarose. Porém, a existência de fases distintas
controladas pelo potássio e pela sacarose ainda não é bem entendida,
necessitando de maiores estudos.
Fatores
ambientais como
a disponibilidade hídrica, temperatura, qualidade e intensidade luminosa e
concentração intracelular de CO2 também controlam a
abertura e o fechamento estomático. O estresse por falta de água nas folhas
leva à síntese de ácido abscísico (ABA) que, chegando às células-guarda,
sinaliza a ocorrência de estresse e induz o fechamento dos estômatos.
O
mecanismo pelo qual o CO2 regula o movimento dos estômatos não
é bem entendido. MANSFIEL et al. (1990) obtiveram resultados opostos:
- as
células-guarda podem ficar mais túrgidas com o aumento da concentração do gás,
favorecendo a formação de malato;
- as
células-guarda podem diminuir o turgor com o aumento da concentração do CO2.
Na
maioria das espécies vegetais, o estômato se abre na presença de luz e fecha no
escuro. Porém, a luz azul (comprimento de onda em 460 nanômetros) estimula a
abertura estomática, independentemente da concentração do CO2; a luz
vermelha (comprimento de onda em 700 nanômetros) estimula a abertura estomática
nos cloroplastos das células-guarda.
Radiação Solar
O Sol é a
fonte universal de energia. Toda a vida em nosso planeta é direta ou
indiretamente dependente da fotossíntese dos organismos clorofilados (exceto
bactérias quimioautotróficas). O oxigênio tornou-se necessário
para a respiração como fonte de energia para o metabolismo e os carboidratos tornaram-se
substratos para a respiração e o ponto de partida para diferentes rotas
biossintéticas.
Cerca de
5% da energia solar que alcança a Terra pode ser convertida em carboidratos
através da fotossíntese. Aproximadamente 44% das radiações que chegam à Terra
são as radiações visíveis utilizadas na fotossíntese. Aqui nos trópicos, a
radiação solar é máxima no verão (dezembro-janeiro) e mínima no inverno (junho-julho).
Podemos
separar a luz em diferentes cores, isto é, em comprimentos de ondas, como sua
passagem através de um prisma, como visualizamos no espectro magnético abaixo:

Como os
tipos de radiações influenciam as moléculas?
A Radiação
Ultra-Violeta promove a ionização de moléculas, ou seja, tem o
comprimento de onda muito curto e o quantum (um fóton carregado de energia) é
muito elevado e consegue comprometer a estrutura de uma molécula orgânica. Já a Radiação
Infra-Vermelha é composta por fótons de comprimento de onda mais
longos, com quantum baixo; ao serem absorvidos alteram a cinética das
moléculas, elevando a temperatura, por exemplo. A Radiação Visível possui
nível energético o suficiente para excitar elétrons entre os orbitais
eletrônicos das moléculas causando reações químicas. Esses três tipos de
radiação são as mais importantes para a fotossíntese.
Mas o que
acontece com a radiação quando incide em uma folha?
A radiação,
nas folhas, pode ter três destinos:
1. Reflexão:
parte da radiação incidente retorna da folha; a capacidade de reflexão da folha
depende da natureza da superfície foliar, como por exemplo, uma folha repleta
de tricomas aumenta a reflexão.
A
radiação visível tem reflexão entre 6 e 10%; a radiação ultravioleta é
fracamente refletida (não mais que 3%) e a radiação infravermelha reflete cerca
de 70% da radiação.
2. Absorção:
cerca de 70% da radiação é absorvida no interior da folha e diminui sua
intensidade ao atravessar as camadas celulares do mesófilo foliar.
A
absorção da radiação visível depende dos pigmentos fotossintetizante (que
veremos na próxima aula). A radiação UV é barrada pela cutícula e pela parede
externa suberizada da epiderme e menos de 1% dessa radiação alcança camadas
mais interiores da folha. O mesmo ocorre com a radiação infravermelha.
3. Transmitância:
depende da estrutura e espessura da folha. Por exemplo, folhas flexíveis
transmitem (ou seja, “deixam passar”) entre 10 e 20% da radiação solar; folhas
finas transmitem até 40%; folhas espessas e duras transmitem abaixo de 3% da
radiação. Os comprimentos de onda mais transmitidos são o verde e o vermelho.
Pigmentos Fotossintetizantes
Qualquer
substância que absorve luz visível é um pigmento. Quando absorvem
luz, os elétrons dos pigmentos são impulsionados a um nível de energia mais
elevado. Quando os elétrons retornam, a energia pode ser dissipada como calor;
pode ser reemitida como energia luminosa, fenômeno conhecido como fluorescência e
a energia pode ser capturada para a formação de ligações químicas.
Os pigmentos
fotossintetizantes, isto é, que participam da fotossíntese, são as clorofilas e
os carotenoides. A Clorofila a, presente em
todos os organismos eucariontes fotossintetizantes e nas cianobactérias, é
constituída por um anel de porfirina e um hidrocarboneto de 20 carbonos chamado fitol.
A Clorofila b é
considerada como um pigmento acessório, encontrada nas plantas, algas verdes e
euglenófitas. A clorofila b difere da clorofila a pela
substituição do grupo metila (-CH3) ligado ao anel da porfirina,
pelo grupo formila (-CHO), como mostra a figura abaixo. Outras clorofilas, como
as do tipo c, d e e também atuam
como pigmentos acessórios.
Os carotenoides são
o segundo grupo de pigmentos mais abundantes nas plantas. Sua estrutura básica
é um esqueleto com 40 átomos de carbono com ligações duplas alternadas,
mostradas na figura abaixo, como exemplos o a e o b-carotenos.
Atuam na absorção de luz, como pigmentos acessórios e nafotoproteção do
aparato fotossintético, pois canalizam a energia em excesso para fora das
clorofilas e desativam os radicais livres.
As xantofilas,
também na figura abaixo, contêm grupos hidroxilas ligados à cadeia carbônica
constituída de 40 carbonos, atuando também como pigmentos acessórios.
Exemplos de xantofilas são a zeaxantina e a luteína (Figura
1).
 |
| Figura 1 – Estrutura molecular de pigmentos fotossintetizantes. Fonte: MAJEROWICZ, 2004, p. 128. |
Aparelho Fotossintético
Na figura 1, visualizaremos as estruturas de um cloroplasto.
Podemos observar que o cloroplasto é composto por uma Membrana externa ou envelope que
define os limites do cloroplasto. É constituída por uma dupla camada que
circunda o sistema interno de membranas e a matriz fluida gelatinosa que
preenche os espaços internos dos cloroplastos, denominada estroma. No estroma há enzimas, DNA, ribossomos e amido
para a manutenção do metabolismo celular.
 |
| Figura 1 – Estrutura de um cloroplasto. Fonte: LOPES, www.editorasaraiva.com.br/biosonialopes (2006). Acesso em: 13/06/2006, modificado. |
As Tilacoides são
membranas duplas com formato de vesículas achatadas e empilhadas que contêm as
clorofilas. As clorofilas agrupadas formam estruturas denominas de complexos
de antena; local onde ocorrem as reações de luz da fotossíntese. O Granum são
tilacoides associadas entre si, formando “pilhas de moedas” (devido ao formato
achatado).
Mas o que
acontece quando os pigmentos absorvem a energia solar?
Primeiramente,
os elétrons são absorvidos pelas clorofilas no complexo antena. As
demais clorofilas e os carotenoides auxiliam na absorção luminosa.
Em
seguida, ocorre excitação do elétron, levando-o de um estado de baixa energia
(não excitado) para um estado de alta energia, como podemos ver na figura 2.
O elétron
excitado relaxa e volta ao primeiro nível do estado excitado. Quando ele
retorna ao estado fundamental, podem ocorrer três processos:
1) perda de calor;
2) emissão de um fóton (fluorescência)
e
3) transferência de
energia a uma molécula adjacente, onde outro elétron é excitado.
 |
| Figura 2 – Estados de excitação do elétron após ser absorvido pela clorofila. Fonte: MAJEROWICZ, 2004, p. 127. |
Parabéns trabalho excelente.
ResponderExcluir